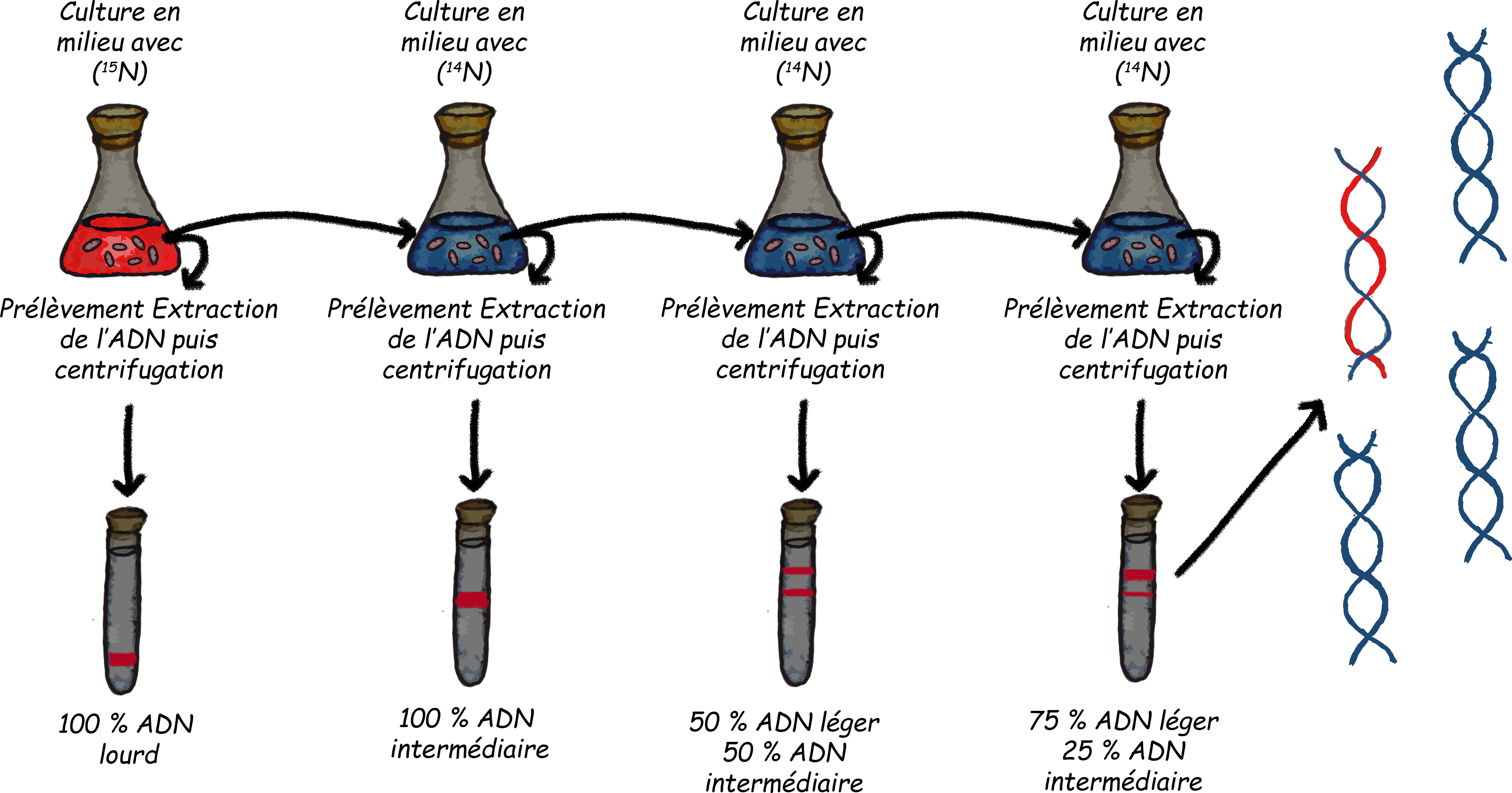
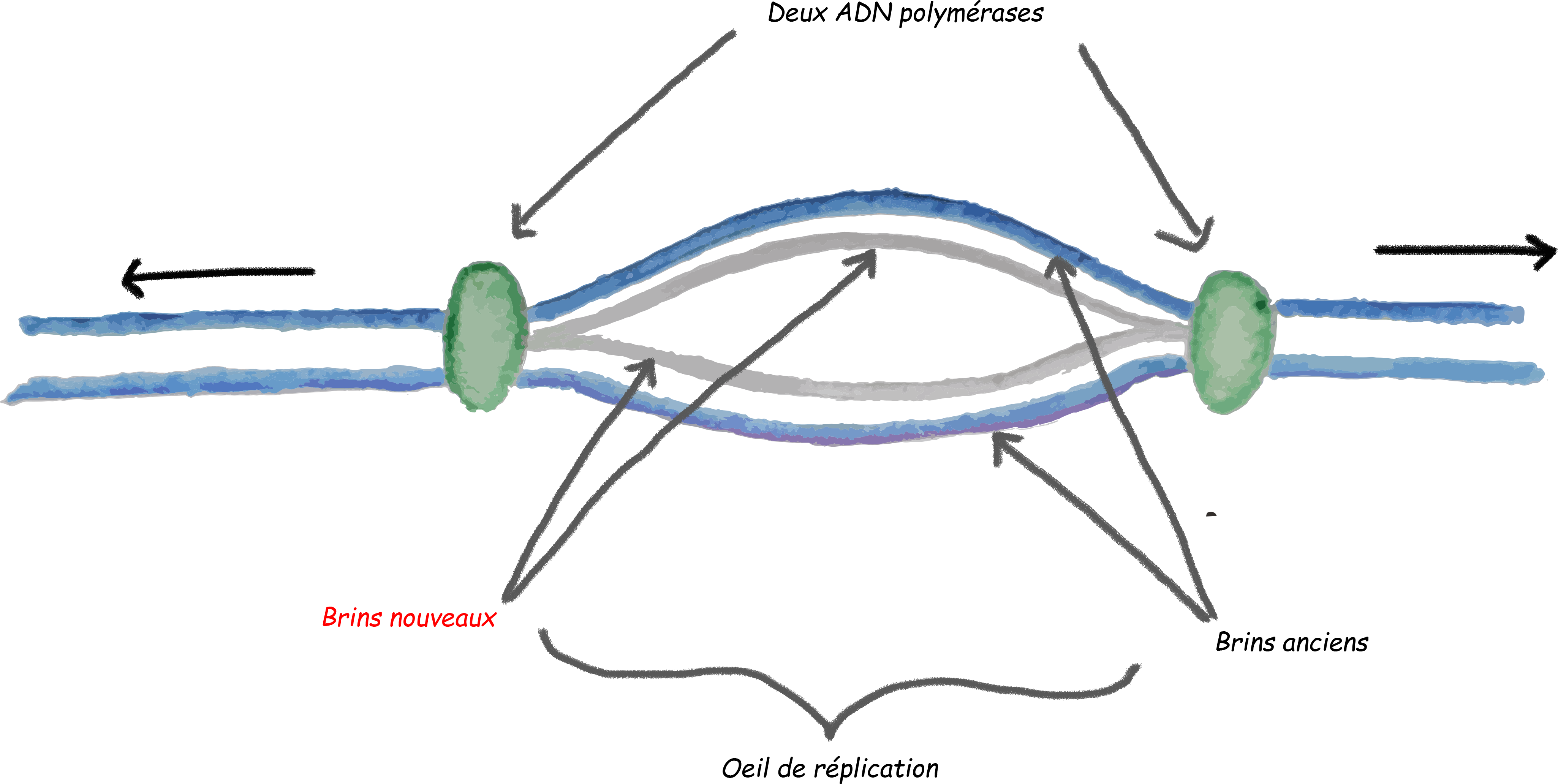
N0
N1= 2N0
N2= 2x2N0=22N0
N3=2x2x2N0=23N0
Donc après n génération Nn=2n N0
Le temps qui sépare deux divisions successives (ou temps nécessaire au doublement d’une population) est appelé temps de génération G.
Il est noté G :
G = temps / nombre de division (en min)
Le taux de croissance horaire (r) lui exprime le nombre de divisions effectuées par unité de temps.
r = n / t ⇒ n = rt (en div/h)
Avec n=rt et N=2n N0
On peut noter : N=2rt N0 (1)
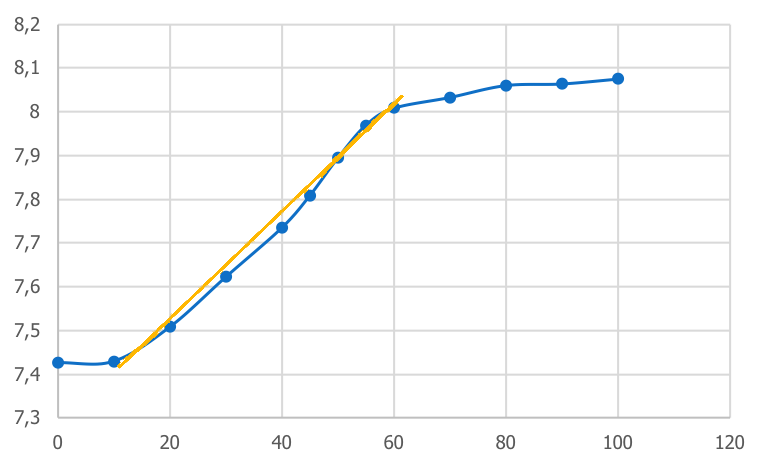
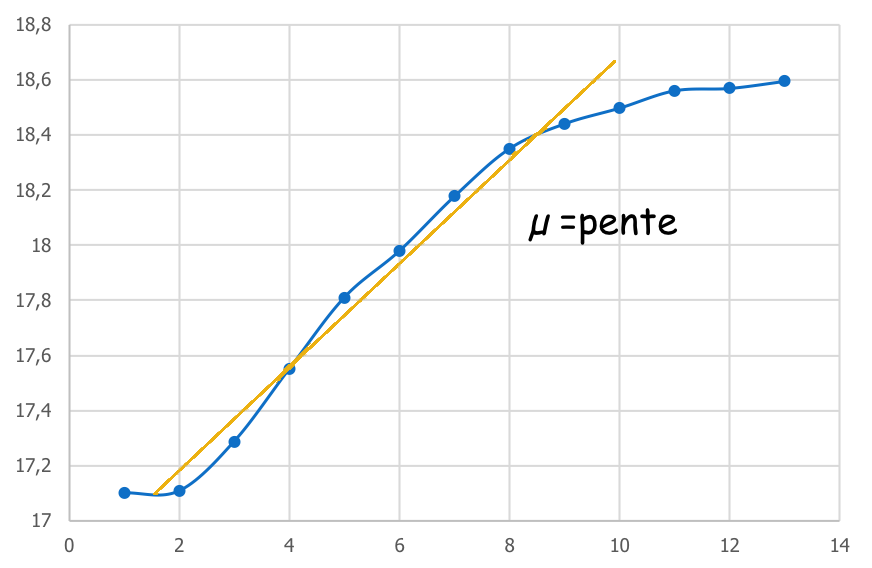
Si l’on repart de notre courbe N=f(t), et que l’on s’intéresse à la partie exponentielle, ou la vitesse de division est constante et maximum, on observe que cette phase « tend » vers la formation d’une droite.